
生物大分子起源的悖论——“鸡”与“蛋”之争
在细胞进化过程中,先有核酸还是先有蛋白?先有复制还是先有代谢?这些依然是生命起源中的未解之谜。在生物个体水平,亦普遍存在类似的问题,如先有‘鸡’还是先有‘蛋’?或是先有‘雌’的还是先有‘雄’的?……这些看似简单的问题,却是现代科学无法解答的悖论,但我们岂可一避了之?
1. 蛋白质与核酸之比较
蛋白质仅由氨基酸通过肽键连接而成,而核酸由核苷酸聚合而成,核酸含有三种组分:核糖(或脱氧核糖)、碱基和磷酸。构成蛋白质的氨基酸有20种(不到天然氨基酸的1/10),构成核酸的核糖只有两种(RNA中为核糖,DNA中为脱氧核糖),核酸中的碱基也不过5种(RNA和DNA各有4种,两者之间仅有一个碱基不同)。
DNA是核苷酸的长链聚合物,通常是两条DNA单链通过碱基之间的氢键形成相互缠绕的双螺旋结构(图1)。两条DNA链遵循所谓“配对”原则,即腺嘌呤(A)与胸腺嘧啶(T)配对,鸟嘌呤(G)与胞嘧啶(C)配对(图3),并且绝不会有例外。与DNA规则的双螺旋结构相比,RNA的结构复杂而多样,它区分为三种不同类型:tRNA、mRNA和rRNA,分别行使不同的功能,也具有十分不同的结构特点(图1)。其实RNA(如mRNA)的结构也颇为复杂,靠随机性产生出这样的分子也是天方夜谭!
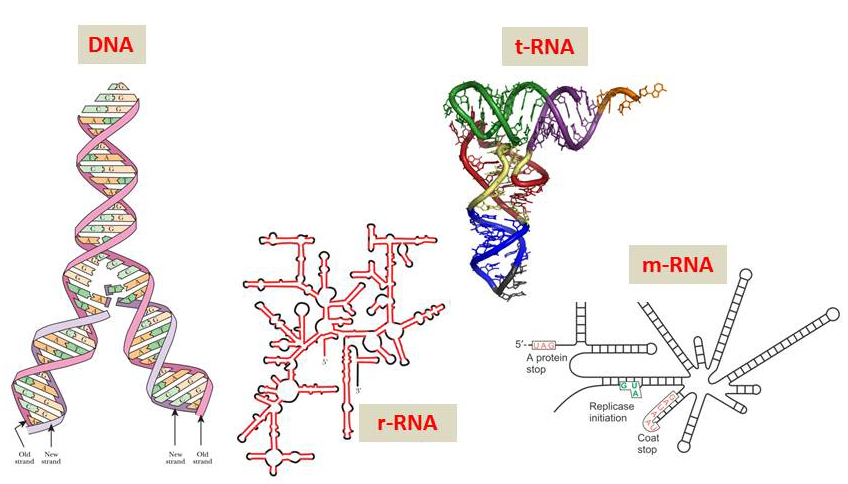
图1 脱氧核糖核酸(DNA双螺旋)和核酸(RNA)(引自Weaver 2008,David 2010)
DNA双螺旋之间的碱基配对规则源自不同结构碱基的契合:4种碱基中,2种为单环,两种为双环,因此只有两种配对方式有利于维持双螺旋结构的稳定性—即A与T、G与C,前者通过两对氢键,而后者通过3对氢键。这样的设计,既能维持DNA的双螺旋结构,又方便复制,也方便转录为RNA(但在RNA中用U取代了DNA中的T)。
与核苷酸相比,氨基酸在种类及结构上(分为1-4级结构)要多样得多,这使得蛋白质能形成各种复杂的立体结构(图2),这是蛋白质(作为酶)具有几乎无限催化可能性的重要结构基础。蛋白质肽链内的氨基酸之间的作用力也十分多样,包括氢键、疏水作用、离子键和二硫键等,它们维持蛋白质的2~4结构。
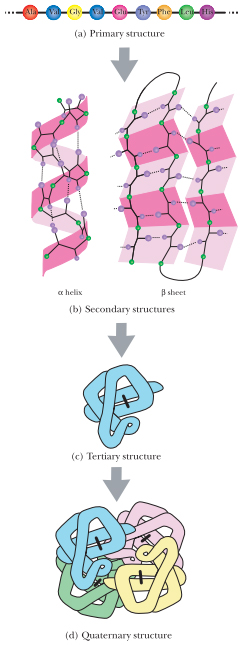
图2 蛋白质的1~4级结构(引自Clark 2010)
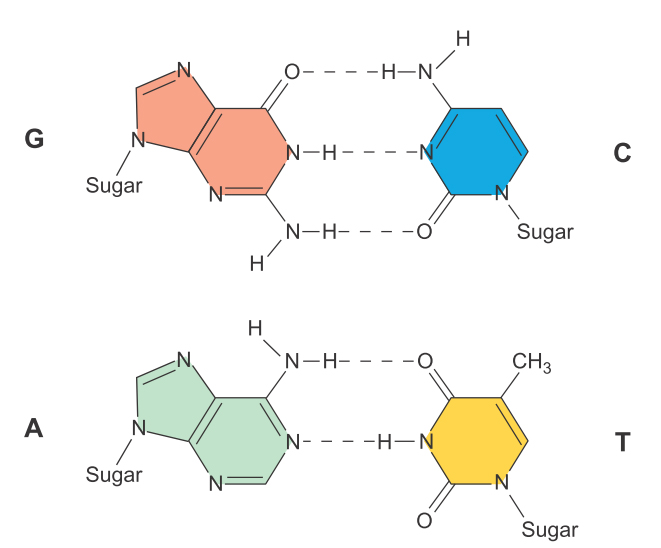
图3 连接在DNA两条磷酸脱氧核糖骨架上的碱基通过氢键彼此联系在一起,配对原则是A-T,G-C(引自Weaver 2008)
2. 先有核酸(信息)还是先有蛋白(结构)?
在现存的任何生命系统中,蛋白质和核酸相互依存,不可割裂,因为如果没有蛋白质的参与,DNA/RNA的复制便无法完成,而没有DNA/RNA携带的遗传信息的指导,蛋白质也无从产生。正如土耳其的神创论者雅荷雅(2003)所说,“DNA只能借助蛋白质的一些酶来复制,但这些酶的合成只能依靠DNA的信息码得以实现。它们相互依存时,要么为了复制必须同时存在,要么其中的一个,在另一个之前被‘创造’”。但蛋白质和核酸之间的这种互为因果的关系是如何演化而成的呢?虽然有一些学说试图解答这一问题,但均无法令人满意。
① 蛋白质在先(proteinsfirst)论
在对核酸还没有太多认识的背景下,一般认为首先由无机物形成氨基酸,再进一步形成多肽与蛋白质,最后发展出原始生命。与其说是蛋白质在先,不如说很少考虑核酸的形成问题,如十九世纪的英国博物学家达尔文和德国哲学家恩格斯,以及二十世纪初的苏联生物化学家奥巴林、美国化学家米勒等。
② 核酸在先(nucleicacids first)论
核酸在先论者认为,首先由核苷酸分子随机聚合成RNA进而出现了核酶—一种同时具备自我复制能力和催化功能的RNA分子。核酶的发现打破了只有蛋白质才是酶的传统观念(Kruger et al.1982,Guerrier-Takada et al. 1983)。当然,与蛋白质催化剂(酶)相比,细胞内的RNA催化剂要少得多。从功能上来说,有些核酶能切割RNA或DNA,有些能转移核苷酸或磷酸,有些能进行水解反应等,在已知的几十种天然RNA催化剂中绝大部分参与RNA的加工和成熟。与蛋白质酶相比,核酶的催化效率较低,因此被认为是较为原始的催化酶。但是,笔者认为,酶的催化效率与原始性之间未必存在因果关系。
核酸在先论者认为,由于催化效率和多样化的选择压力可能导致了核酶催化肽基转移(因此形成小蛋白),而与RNA耦合的寡肽复合物则形成了更好的催化剂。最初的核糖体可能就是这样形成的,从而导致了更普遍的蛋白质合成方式的出现。后来,被合成的蛋白质可能在催化能力上比核酶更强,因此,变成了占优势的生物大分子,使核酸降格到现代的用处,即占绝对优势的功能就是作为遗传信息的携带者。但笔者要问的是,为何出现催化效率和多样化的选择压力?为何会出现遗传信息呢?
另外,有研究显示,一些氨基酸与它们相对应的密码子有选择性的化学结合力,这显示现在复杂的蛋白质制造过程可能并非一早存在,最初的蛋白质可能是直接在核酸上形成(Knight & Landweber 1998)。
③ 不分先后论
德国化学家和生物物理学家、诺贝尔化学奖得主艾根(ManfredEigen)在“The Hypercycle: A principle of naturalself-organization”一书中指出,在生物信息起源上的这种“在先”,不是指时间顺序,而是指因果关系,这里有一种双向的因果关系,或者说是一种互为因果的封闭圈。核酸和蛋白质的相互作用,相当于“封闭圈”即“循环”的一个复杂的等级组织。从反应循环,到催化循环,再到超循环就构成了一个从低级到高级的循环组织(Eigen and Schuster, 1979;中译本:艾根和舒斯特1990)。这个所谓的超循环理论是独创的还是模仿的呢?

首先,古希腊哲学家亚里士多德(Aristotle)曾对循环给予了精辟的阐述,他在《论生成与消灭》中说,“如果事物的生存是必然的,它的生存就是永恒的,而如果是永恒的,也就是必然的。因此,如果某物的生存是纯粹必然的,它就必定是循环的,即返回自身。因为生成必然要么有限度,要么无限度;如果无限度,它必定要么是直线式,要么是圆圈式。但是,如果生成是永恒的,就不可能是直线式,因为它没有开端,无论我们向下考虑(当作将发生的事)还是向上追溯(当作已出现过的事)。但生成必然有开端,虽然不是有限的,而是永恒的。因此它必然是圆圈式……正是圆圈式的运动和生成中,才有纯粹的必然,也就是说,如果过程是圆圈式,其中的每一个环节必然会生成或已经生成,如果过程是必然的,它们的生成就是圆圈式”(亚里士多德1991)。

Aristotle(约公元前384年-公元前322年)
德国哲学家黑格尔(1770年-1831年)曾论述过认识的无限循环过程,他在《逻辑学》一书中在解释哲学开端时指出,“开端的规定性,是一般直接的和抽象的东西,它的这种片面性,由于前进而失去了;开端将成为有中介的东西,于是科学向前运动的路线,便因此而成为了一个圆圈……那个造成开端的东西,因为它在那里还是未发展的、无内容的东西,在开端中将不会被真正认识到,只有在完全发展了的科学中,才有对它的完成了的、有内容的认识,并且那才是真正有了根据的认识”(黑格尔1922)。在这里,黑格尔似乎是借用了赫拉克利特关于“在圆周上,起点和终点是同一的”的说法,并将其引申到了认识过程。
此外,黑格尔在他的《自然哲学》一书中指出:“个体性把自身规定为特殊性或有限性,而又否定这种有限性,并向自身回归,在过程的终点使自己恢复为开端,这是一个无限的过程”(黑格尔1986)。
德国哲学家康德(KantI,1724年-1804年)在1790年出版的专着《判断力批判》中指出,“是自然目的的东西就是有机体”,而“如果一个事物同时是原因而又是它自己的结果(即令是在双重的意义上),它就是作为一种自然目的的”。
以笔者之见,艾根对所谓“超循环”的描述真切地混杂了亚里士多德、黑格尔和康德的思想,说模仿了哲学家的口吻一点都不为过,当然身为德国科学家接受过自己祖国的伟大哲学家的思想熏陶也不奇怪。艾根试图借用哲学家的智慧来揭示一个世纪难题,但遗憾的是,他的整合只是个半吊子!他牵强地将这一问题与细胞内的各种循环扯上了关系,认为他的超循环具备内在的整合信息能力。但问题是,这种整合的动因到底是什么呢?
3. 先有复制(基因)还是先有代谢?
复制优先论者认为,核酶及其互补链可以自我复制并通过突变产生新的代谢反应,而相比之下,代谢似乎并不具备进化的能力,代谢循环中的要素是否能够自我组织是值得怀疑的,而自动催化循环通常都有内在的困难,因此,由于代谢自行进化的这些问题,复制应该在代谢之前(Orgel 2000,2008,王子晖2011a)。
但是,威廉斯(2001)认为,“DNA分子的极端特化迫使我们认为核基因是一个后来发展的产物。早期遗传系统的建立也是极不可能的”。
代谢优先论者则认为,一套燃烧热很低的小分子水溶性有机物就构成了三羧酸(TCA)循环中所有的有机酸,这个发现非常有启发性,它意味着这些有机酸可能自发组织行成了还原性的TCA循环。这样,前生物进化的第一步就可能是通过分子间的联系进行的。相反,若RNA复制子随机产生序列,那么这些序列可能没有一点用处(Morowitz et al. 2000,王子晖2011a)。
Robinson(2005)评价道,“这两个模型—通常被称为‘基因优先论’和‘代谢优先论’—都仍有显着的问题,并且没有哪一个是完善或具有明显优势”。王子晖(2011a)指出,“若代谢不能有效引导复制,复制又不能有效引导代谢,那么这个两难处境将只能有一种解决方式:它们必须从一开始就共同作用。任何一种合作要想成功,每一方必须对另一方有所帮助。复制子可以通过引发催化新的代谢反应参与代谢。而代谢是如何帮助复制的呢?由于复制面临的最大挑战是需要从无数没用的序列中挑选出有意义的那些,代谢反应能够对此提供帮助。复制子可以帮助产生新的催化类型,而代谢作用可以帮助选择有用的复制子,它们的共同作用便可以迅速发展出一个基于可用结构单元和能量的生命策略蓝图,在不违背基本热力学规律和水溶性的前提下,建立由代谢中固有的分子逻辑构成的代谢通路、循环和网络”。
笔者认为,这样的观点似乎又是仿效了艾根关于核酸(信息)与蛋白质(结构)之间关系的说法,基因与代谢之间原则上可能存在一种不可分割的协同演化关系,但问题是,到底是何种缘由引发了这样的进化过程呢?或者说,核酸和蛋白为何要协同演化呢?
4. 为何生命只有一套遗传密码?
一个DNA链可能包含数百万个相连的核苷酸,但碱基却只有A、T、G、C四种。遗传密码使用的是一种4进制代码,就是由DNA链上这4个碱基的排列顺序所决定。构成RNA的碱基有四种,由于一个氨基酸由三个碱基所决定,这样从理论上分析碱基的组合有43=64种,但有3种终止密码子,并不编码氨基酸,因此,有61个编码氨基酸的密码子。但是,构成蛋白质的氨基酸只有20种,这就意味着存在两个或多个密码子编码同一个氨基酸,这称为简并性,已知大部分密码子具有这种特性。
英国进化生物学家道金斯(RichardDawkins)将形形色色的生命比作运载遗传指令的工具,而制造这些工具的方法就是DNA的序列,他在“The Selfish Gene”(译文:自私的基因)一书中感叹道,“工具本身千变万化,但是,相反地,制造这些工具的方法却极其单一,令人难以置信:仅仅是A、T、C、G的排列一次又一次地突变”(道金斯2008)。
艾根问道,动植物的种类有数百万种之多,而细胞却只有一种基本的分子机制,即一种普适性的遗传密码,这是为什么?他认为,生物的遗传密码也是在分子进化阶段确立起来的。他说,“在达尔文物种进化的前面,还有一个类似的分子进化的渐进过程,由此导致了唯一的一种运用普适密码的细胞机制。这种密码最终确立起来,并不是因为它是唯一的选择,而是由于一种特殊的‘一旦—永存’选择机制,这可以从任何随机分配开始”(艾根和舒斯特1990)。
我要问,什么叫做‘一旦—永存’的选择机制?“一旦”都“永存”了,还选择什么?这不是自相矛盾的逻辑吗?
一般认为,原始的遗传密码可能比今天简单得多,随着生命演化制造出新的氨基酸再被利用而令遗传密码变得复杂。但是,这又是如何才能实现的呢?在最初生命处于无性生殖的状态下,现在生物界的唯一一套统一的密码要么意味着它在生命起源的第一个物种那里就得到了完美的发展,然后才开始物种的分化,要么它在物种间能够不断交流。到底是什么?
结语
生命科学中的许多“鸡”与“蛋”(蛋白质与核酸、雌与雄等)之争,既是悖论,又是世纪之谜!这样的悖论又交织于一种自然的目的性之中,正如德国哲学家康德所言,“在像这样的一个自然产物里,我们是把每一部分都认为是由一切其余的部分的作用而由其存在的,而且又是为了其他各部分以及为了全部而存在的……这个部分必须是产生其他部分的一个机官—所以每一部分都是交互产生其他部分的……只有在这些条件下而且按这些规定,一个产物才能是一个有组织的并且是自组织的物,而作为这样的物,才称为一个自然目的”。问题是,这个自然目的到底意指何物呢?它是如何牵引了这种“鸡”与“蛋”的演化之旅的呢?(bio.angelyeast.com)



